Felipe da Silva Lima, Caroline Dutra Lacerda y Hernan Šimovic
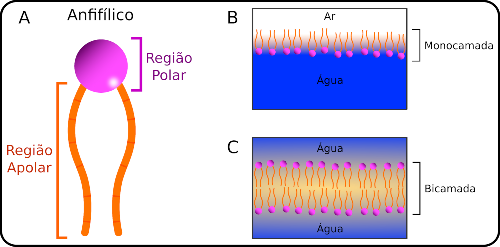
Las membranas biológicas (MB), las estructuras esenciales para sustentar la vida, separan el entorno interno (el citoplasma) del entorno externo y definen los compartimentos dentro de la célula. Los MB están compuestos principalmente de lípidos y moléculas anfifílicas que constan de una región polar (soluble en agua) y una región no polar (también llamada hidrófoba o insoluble en agua) (Fig. 1a). Otras moléculas, como proteínas, carbohidratos e incluso ARN, pueden formar parte de los MB.
forma 1 شكل
La idea de que la célula es una solución delimitada por una barrera proviene de la observación de que las células responden a la tensión del medio, lo que indica la presencia de una membrana semipermeable. Los modelos de membranas para el estudio de la ósmosis, término introducido por el químico T. Graham en 1854, un tema muy popular en ese momento, ayudaron a apoyar esta idea, que se reforzó al mostrar la asimetría en la distribución iónica dentro y fuera de la célula. La relación entre la aversión a los solutos y su absorción por los tejidos vivos llevó a CE Overton, al final del noveno hexágono, a sugerir que la membrana celular tendría una naturaleza lipídica.
Así, a principios de sXX, se aceptaron dos conceptos básicos de las propiedades de esta membrana: un límite físico de naturaleza lipídica que presenta respuestas osmóticas. La forma utilizada hoy para describir megabytes es mosaico fluido. En este modelo, los lípidos forman una estructura bicapa, las porciones polares están expuestas al agua tanto por dentro como por fuera, y las regiones no polares forman una región hidrófoba. Esta orientación específica de los lípidos en la bicapa es impulsada por el llamado efecto cardiogénico del agua, que mejora la separación de agua y aceite. Todavía en mosaico fluidoLas proteínas y los carbohidratos se distribuyen en este «mar» de grasas. Y precisamente porque este modelo es aceptado hoy en día y se usa incluso en los cursos de biología de la escuela secundaria, comprender su origen, así como la evolución de sus supuestos, es lo mejor para nosotros.
Nuestra historia comienza con el estudio de sistemas sencillos a base de petróleo en la zona ribereña. En 1774, Benjamin Franklin publicó un artículo en el que describía cómo las gotas de petróleo frenarían la formación de olas en la superficie del agua; los marineros de algunas partes de Europa tenían la superstición de verter petróleo en el agua para «calmar el océano». Franklin también notó que las gotas de aceite agregadas a la superficie del agua se difunden y forman películas muy delgadas, mientras que las gotas de aceite agregadas a superficies duras como el mármol siguen siendo gotas con poca deformación.
En 1890, Lord Rayleigh, utilizando un contenedor de área específica, determinó que el espesor de la capa de aceite necesaria para cubrir toda la superficie, sin exceso o escasez de material, era de 1,63 nanómetros (nm), casi 43.000 veces menor que el espesor de ¡Un pelo! Esta medición ha sido mejorada en gran medida por Agnes Pockels, una científica autodidacta que, por sí sola, desarrolló un dispositivo experimental que permite medir el espesor de la película de aceite con mucha mayor precisión. Pockels corrigió la medición de Rayleigh (1,30 nm) y también concluyó que el espesor de la monocapa depende del tipo de aceite utilizado.
El siguiente gran salto en esta historia se produjo en 1917, cuando Irving Langmuir mejoró el sistema que Buckles había desarrollado y utilizado para estudiar moléculas anfibias en la interfaz del agua. Langmuir avanza en la interpretación de los datos, señalando que las capas formadas eran «monomoléculas» (capas tan gruesas como una sola molécula), un término ahora conocido como monocapas. Además, concluye que las moléculas anfifílicas se orientan preferentemente a sí mismas en la interfaz, con los grupos polares enfrentados al agua y los grupos no polares expuestos al aire (Fig. 1b). Según Langmuir, esta disposición era el resultado de fuertes interacciones entre el agua y los grupos polares de moléculas anfifílicas, lo que llevaría a los grupos no polares de moléculas anfifílicas a permanecer en un área libre de agua, expuesta al aire. La orientación preferencial de las moléculas anfifílicas en la interfaz aire / agua se debería al efecto hidrofóbico.
Los resultados de Rayleigh, Pockels y Langmuir, considerados como pertenecientes a la química física, no recibieron mucha atención de la comunidad biológica hasta 1925. El pediatra Evert Gorter pensó que los experimentos físicos y químicos podrían usarse para determinar la estructura de las membranas biológicas. En ese momento, se sabía que las membranas estaban compuestas de lípidos, pero se desconocía la estructura de las membranas.
Gorter y su estudiante de posgrado Francois Grendel extrajeron lípidos de los glóbulos rojos de diferentes animales y, utilizando el dispositivo experimental desarrollado por Pockels y mejorado por Langmuir, determinaron el área ocupada por una monocapa de estos lípidos. Dado que se conocía el área de superficie de los glóbulos rojos, encontraron que el área ocupada por la grasa era dos veces más grande que el área de superficie de los glóbulos rojos. Concluyeron que los glóbulos rojos deben estar cubiertos con dos capas de grasa. Dado que Langmuir ya había determinado que las regiones no polares de los lípidos no entran en contacto con el agua en las monocapas, Gortel y Grendel sugirieron que las dos monocapas en las membranas se organizarían de tal manera que sus regiones polares estarían expuestas al agua (interno o medio externo). de la celda), con las regiones no polares orientadas hacia el centro. Así, el concepto de MB nació de la bicapa lipídica (Fig. 1C).
Bangham y Horn validaron este concepto utilizando microscopía electrónica al observar la similitud entre el patrón de tres capas de la membrana externa de la célula y las estructuras compuestas de lecitina (una mezcla que consiste principalmente en fosfolípidos extraídos de la yema de huevo) dispersos en exceso de agua.
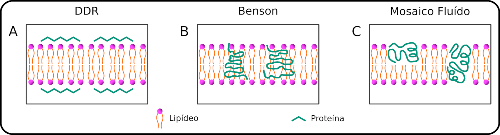
En 1935, Danieli y Davson sugirieron, utilizando argumentos termodinámicos y medidas de tensión superficial y permeabilidad, que en la bicapa lipídica propuesta por Gorter y Grendel, también debería haber una película de proteínas globulares adsorbidas en la interfaz. Los autores llamaron a este modelo un «modelo de mosaico», en el que las proteínas y los lípidos son rígidos con poco o ningún movimiento molecular. Este modelo fue modificado por J. Robertson para acomodar proteínas no esféricas y adaptó el modelo a los resultados de difracción de rayos X. Las proteínas todavía estaban en el exterior de la membrana celular. Este modelo de MB (DDR, Figura 2a) fue aceptado hasta principios de la década de 1970.
Aquí, notamos el peso conceptual de los modelos aceptados, ya que en el modelo DDR, tanto las proteínas como los lípidos ocupaban lugares fijos y el sistema era escasamente móvil. Es interesante (pero quizás no muy útil) preguntar cuánto se evita la inversión de este modelo estático y los experimentos que pueden mostrar, mucho antes, la dinámica de la membrana.
A mediados de la década de 1960, las imágenes de microscopía electrónica ya mostraban que algunas proteínas estaban ancladas en la bicapa lipídica y no adsorbidas en la interfaz, como sugería el modelo DDR. Al mismo tiempo, Benson y Green demostraron que los fragmentos que contienen proteínas y algunos lípidos pueden separarse de las membranas internas de las mitocondrias, y que estos fragmentos que se reconstruyen en las membranas lipídicas recuperan su actividad. Sobre la base de estos datos, Benson propuso un modelo (Fig. 2b) en el que algunas proteínas se unen estrechamente a los lípidos y se insertan en las bicapas lipídicas, que permanecen rígidas.
A principios de la década de 1970, se discutieron dos modelos distintos de MB (Benson y DDR). Ambos explican solo algunas de las propiedades de las membranas. Por ejemplo, en 1968, Luzzati demostró que las cadenas no polares de lípidos de membrana se parecen más a un líquido (alta movilidad) que a un sólido. Esta observación no se explica por ninguno de los modelos, en los que los lípidos de la bicapa son sólidos.
En 1972, Singer y Nicholson publicaron un artículo titulado «Modelo de mosaico fluido de la estructura de las membranas celulares» o «Modelo de mosaico fluido de la estructura de las membranas celulares». Una vez más, el modelo se basa en el efecto catastrófico del agua y en varias observaciones experimentales, incluidas medidas importantes del movimiento de lípidos y proteínas en las bicapas. Los resultados de la navegación llevaron a Singer y Nicolson a llamar al modelo un «mosaico fluido», en contraste con las estructuras sólidas propuestas anteriormente. En el modelo de mosaico fluido, los lípidos pueden moverse libremente, formando un «solvente» en el que las proteínas pueden anclarse (Fig. 2c). Sin embargo, los lípidos no deben considerarse adyuvantes en este modelo, argumentan los propios autores, porque las interacciones de lipoproteínas específicas son esenciales para mantener las funciones de las proteínas de membrana. Prueba de ello es la gran variedad de lípidos presentes que pueden formar membranas y las diferentes proporciones de lípidos y proteínas que se pueden encontrar en las células. A pesar de esto, la gran mayoría de las membranas celulares deben compartir las propiedades descritas (y esperadas) por el modelo de mosaico de fluidos. Numerosas observaciones han apoyado (y apoyado) el modelo, y su capacidad predictiva ha llevado al abandono de otros modelos.Hoy en día, el modelo de mosaico fluido es la vista canónica de la estructura de la membrana.
El debate sobre el modelo canónico queda para otro artículo, ya que la evidencia más reciente trae a la pantalla la generalidad del modelo de mosaico fluido.
Felipe da Silva Lima es profesor de la Universidad Federal de Pernambuco (UFPE) y Caroline Dutra Lacerda y Hernan Chaimowicz son profesores de la Universidad de São Paulo (USP)